~~~~~~~~~~~~~~~~~~~~~~~~~~ 感覺
维基百科,自由的百科全书
感覺 是對客觀現實個別特性(聲音、顏色、氣味等)的反映,由來自物質世界的一定刺激直接作用於有機體的一定感覺器官 ,如光線引起視覺,聲波引起聽覺;刺激在感官內引起的神經衝動,由感覺神經傳導於大腦皮層 的一定部位產生感覺。
感覺是感官、腦 的相應部位和介於其間的神經三部分所聯成的分析器統一活動的結果;無機界沒有感覺,只有跟感覺類似的特性,即單純的物理或化學反映;隨著生命出現,產生了生物反映模式,即刺激感應性;刺激感應性已包括感覺的萌芽;正是在刺激感應性基礎上發展起來的感覺;動物感覺能力在進化 中隨分析器的專門化發展;人類的感覺在複雜的生活條件下和變革現實活動中得到高度發展;人與動物的感覺不同,動物的感覺只是自然發展的結果,人的感覺則包括社會 發展的產物。
感覺屬於認識的感性階段,是一切知識的源泉;它同知覺 緊密結合,為思維 活動提供材料;感覺因分析器的不同分為視覺 、聽覺 、味覺 、嗅覺 、膚覺、運動覺、機體覺、平衡覺等。
神經系統
维基百科,自由的百科全书
神經系統 是由神經元 這種特化細胞的網路所構成的器官系統 ,調節動物的動作與在其身體的不同部位間傳遞訊號。動物體藉神經系統和內分泌系統 的 作用來應付環境的變化。動物的神經系統控制著肌肉的活動,協調各個組織和器官,建立和接受外來情報,並進行協調。神經系統是動物體最重要的連絡和控制系 統,它能測知環境的變化,決定如何應付,並指示身體做出適當的反應,使動物體內能進行快速、短暫的訊息傳達來保護自己和生存。
[编辑 ] 組成 神經系統主要由以下兩種細胞組成:
神經元 :神經系統的主要細胞。神經膠質 :神經系統的次要細胞結構與營養。[编辑 ] 興奮的傳輸 在神經系統中的迅速神經信號傳輸主要通過以下兩種方式 :
在神經細胞里:興奮 通過神經纖維 上的電位差傳輸。 在神經細胞間:興奮依靠突觸 前膜釋放的遞質 傳遞到下一個神經細胞的突觸後膜。 [编辑 ] 神經的功能 神經三大主要功能:
感覺功能:身體的內在感覺受器探測如血的酸度,血壓 等內在刺激,在外感覺受器傳送由皮膚等身體末端所接受到的外來刺激情報。這些情報經由感覺神經傳遞至中樞神經。 綜合及指令功能:對於感覺受器所送來的情報進行分析 、整理、判斷,並做出適當的決定。 運動功能:將整理之後的情報,經由運動神經傳遞至末梢,並執行決定。 在1~3的功能當中,中樞神經負責2的功能,周圍神經則負責1和3的功能。末梢神經中的1,稱之為感覺神經或知覺神經(sensory neurons)、3稱之為運動神經(motor neurons)。
[编辑 ] 人的神經系統 人體的神經系統,包括腦 、脊髓 和神經。神經系統只佔人體體重的約3%,然而卻是人體最複雜的系統。 神經系統被分為二部分:
人類也和其他動物一樣,對於體內和體外的環境變化以及壓力,需要一個調節器官來與其緊密聯繫,保持穩定的狀態(恆定性 ),神經系統也和內分泌系統就扮演了非常重要的角色。藉由複雜的神經纖維和其他細胞組織連結這兩個神經系統,人類才能夠因應外界的環境變化而產生適當的身體反應,並且有思考 、記憶 、情緒 變化的能力。
[编辑 ] 中樞神經系統 中樞神經系統(Central Nervous System)組成了神經系統的主要部分,由腦和脊髓構成。
中樞 腦 前腦 大腦 嗅腦 , 杏仁體 , 海馬體 , 新皮質 , 側腦室
間腦 上丘腦 , 視丘 , 下丘腦 , 底丘腦 , 腦下垂體 , 松果體 , 第三腦室
腦幹 中腦 中腦頂蓋 , 大腦腳 , 腦蓋前部 , 大腦導水管
後腦 後腦 橋腦 , 小腦 ,
末腦 延髓 脊髓
[编辑 ] 周圍神經系統 周圍神經系統(Peripheral Nervous System)是指在中樞神經以外的神經纖維。解剖人體後,人類的肉眼可以看得到由許多條神經纖維,外部包以神經膜(neurilemma)而成的神經。 其主要功能是將感官接受之興奮傳至中樞神經系統,又將來自後者之訊息傳至骨骼肌,以使身體運動。
周圍神經系統因分布的部位不同而有三種(有寫教科書和學者只把周圍神經系統的分法歸為二種,這二類就是軀體神經系統和自主神經系統。這寫教科書或學者通常把腸神經系統納入自主神經系統的一部分。不管何類分法,在醫學上都普遍被接受。):
分布在體軀的稱為軀體神經系統 (Somatic Nervous System) 而分布在內臟者稱為自主神經系統 (Autonomic Nervous System) 分布在腸間的腸神經系統 (Enteric Nervous System)(腸神經系統也被稱為腸間神經叢) [编辑 ] 其他生物的神經系統 動物界裡的神經系統因動物而異。刺胞動物 ,例如水螅 ,還有海葵 和水母 ,它們擁有一套脈絡狀的神經網,當其中的一處產生興奮,就會像波浪一樣波及整個神經系統。這些波同時執行傳入和傳出的功能,即不單傳遞有關化學或機械方面的信息,還反饋輸出動物的反射應答。而扁形動物門 動物有著和傳統神經系統更多相似之處,但缺少大腦。環節動物 和被囊動物 則有大腦的雛形,被稱為神經節 ,是神經的束狀集合。被囊動物的神經節和脊椎動物胚胎髮育過程中大腦的發育有相似之處,被認為是脊椎動物腦幹 的雛形。
[编辑 ] 外部連結
難經
维基百科,自由的百科全书
《難經 》是中國古代醫學著作《黃帝八十一難經 》的簡稱,共三卷(亦有分五卷的)。《難經》書名的含義,有二種解釋:以難字作為問難,另以難字做為難易來解讀。難,讀音為「ㄋㄢˋ(nàn)」。《難經》是闡發《黃帝內經 》的疑難和要旨的第一部書。後世將其列為中醫四大經典 之一。
[编辑 ] 作者 作者及成書年代不詳。《史記·卷105·扁鵲倉公列傳 》及《漢書·藝文志》均未載本書。《傷寒雜病論》及《隋書·經籍志》雖提及本書,但未言作者姓名,直至唐楊玄操《難經注》和《舊唐書·經籍志》才提出本書作者為戰國 時秦越人 (扁鵲 )。從書的內容看,成書當在東漢 以前,大約撰於西漢 時。[1]
[编辑 ] 內容概要 本經以分篇方式共計八十一篇稱八十一難 ,約一萬二千字。以闡述中國醫藥基礎理論為主。可分為六大主議題依序為:
脈學:一至二十二難 經絡:二十三至二十九難 臟腑:三十至四十七難 疾病:四十八至六十一難 腧穴 :六十二至六十八難針法:六十九至八十一難 難經在《內經》的基礎上有所發展,尤其是「獨取寸口」的脈診 法、對經絡和臟腑中命門 、三焦 、腎間動氣、奇經八脈 等的論述對後世影響極大。
[编辑 ] 參考文獻 ^ 甄志亞 主編,上海科學技術出版社 ,1997年6月第2版《難經經釋》 ·清·丁錦 著《難經正義》 ·清·葉霖 著
松果體
维基百科,自由的百科全书
松果體 (又叫做松果腺 、腦上體 或第三隻眼 脊椎動物 腦 中的小內分泌 腺體。它負責製造褪黑素 ,一種會對醒睡模式與(季節性)晝夜節律 (en:Circadian rhythm )功能的調節產生影響的激素 [1] [2] 大腦半球 (en:Cerebral hemisphere )之間,被裹在兩個圓形的丘腦 的接合處。
[编辑 ] 位置 松果體是略帶些紅的灰白色,大小約一顆稻穀那麼大(5-8 mm),就座落在上丘 (en:superior colliculus )的上面、髓紋 (stria medullaris )的後下方及在其側面的丘腦 之間。是上丘腦 (en:epithalamus )的一部份。
松果體是一個中線結構,而且由於其常常石灰化 (en:calcification )的關係,所以時常能在顱骨 的X光照 中看到它
[编辑 ] 結構與組成 松果體在人體由被結締組織 間隙所包圍的小葉狀松果腺細胞 (en:pinealocytes )實質 (en:Parenchyma )所組成。而松果體的表面則被軟膜所覆蓋著。
松果腺主要由松果腺細胞 與其他已確認的四種細胞 所組成。
細胞型式 描述 松果腺細胞 松果腺細胞們是由一種有著四到六個突起浮現著的細胞體所組成的。它們製造並分泌褪黑素 。松果腺細胞可以用特殊的注銀方法染色,以便在顯微鏡下觀察。 間質細胞 間質細胞位於松果腺細胞之間。 血管周圍的 噬菌細胞 在松果體中有許多的微血管 ,而血管周圍的噬菌細胞便位在這些血管附近。血管周圍的噬菌細胞是抗原呈現細胞 。 松果體神經元 在高等的脊椎動物之中有神經元分布於松果體內。然而在囓齒目 的動物上沒有這種現象。 肽 能似神經元細胞在一些物種中,似神經元的肽能細胞存在於松果體中。這些細胞可能有旁泌性的控制功能。
松果體受到來自頸上神經節 的交感神經 支配。然而,也受到來自翼顎神經節 與耳神經節 的副交感神經 的支配。甚至有一些神經纖維經由松果體柄穿入了松果體內(主要的神經支配)。最後,在三叉神經節 的神經元則以含有神經肽 類物質「腦垂腺苷酸環化酶活化肽 」(PACAP)的神經纖維來支配松果體。
人類的松果體小囊包含了一種大量變異的沙礫狀物質,名為腦沙 (corpora arenacea,或名為「acervuli」、「brain sand」)。化學分析顯示腦沙是由磷酸鈣 、碳酸鈣 、磷酸鎂 與磷酸銨 所組成。[3] [4] [5] 氟化物 [6]
[编辑 ] 多物種的解剖學 松果腺細胞 在很多非哺乳類的脊椎動物 中非常地像眼 睛的感光細胞 。一些演化生物學家相信脊椎動物的松果體細胞與視網膜細胞共有一個同樣的演化 原型。[7]
在一些脊椎動物中,曝曬在光線下可以啟動在松果體內的酵素連鎖反應以校正晝夜節律 。[8] 孔 。這與生理學中例如像七鰓鰻 和喙頭蜥 的現代「活化石」以及一些其他的脊椎動物所擁有的顱頂眼 或所謂的「第三隻眼」有關,而在這些動物之中的一部份的顱頂眼具有感光功能。第三隻眼的存在代表著演化早期的視覺感受途徑。[9] 喙頭蜥 內的第三隻眼結構與角膜 、晶狀體 和視網膜 類似,雖然其結構相似度還比較接近章魚而非脊椎動物的視網膜。其不對稱的整體由偏向左邊的「眼」及偏向右邊的松果體囊組成。「在含哺乳動物在內的失去顱頂眼的動物,松果囊被保留下來並壓縮成松果體的形式。」[9]
不像許多哺乳動物大腦的其餘部分,松果體並未被血腦障壁 系統所隔離[10] 腎 。[6]
化石很少保留軟質的解剖結構。大約已有九千萬年歷史的俄羅斯梅羅瓦卡(Melovatka)鳥的大腦是個例外,並展現了一個遠超過預料之外的顱頂眼與松果體。[11]
在人類與其他的哺乳動物之中,從眼睛經由視網膜下視丘路徑 (en:retinohypothalamic tract )系統到視叉上核 (en:suprachiasmatic nucleus )與松果體的光線訊號對於晝夜節律 是必須的。
[编辑 ] 功能 松果體最初被認為是某個較大的器官退化 的殘留物。在1917年早期時,母牛的松果體萃取物被用來使青蛙的皮膚變亮。皮膚醫學教授艾倫‧本生‧勒納 (Aaron B. Lerner)與他在耶魯大學 的同事,在1958年時離析並命名了褪黑素 ,並希望這個來自松果體的物質能夠治療皮膚病。[12] 大鼠 松果體的移除加快了其卵巢的成長、讓大鼠保持在不斷的日光下會減少他們松果體的重量以及松果體切除術(pinealectomy)和持續的日光兩者都對卵巢的成長有同樣程度的影響等不可思議的事實,這些知識促使了名為「時間生物學 」的新領域出現。[13]
褪黑素是色胺酸 這種胺基酸 的衍生物,在中樞神經系統 裡還有其他的功能。黑暗會刺激松果體對於褪黑素的製造,反之光亮則會對其抑制。[14] 視叉上核 後,便產生了自然的晝夜節律。其過程是經從視叉上核投射到室旁核 (paraventricular nuclei)的神經纖維傳達生理節奏的訊號到脊髓 並經交感神經系統到頸上神經節 (superior cervical ganglia),又從那裏傳到松果體。對於褪黑素在人體中的功能依然不太清楚,不過一般將它作為晝夜節律性睡眠障礙 (circadian rhythm sleep disorder)的配藥。
化合物松香烴 (pinoline)亦是在松果體製造。它也是β-咔啉 (β- carboline )之一。
人類的松果體會成長到大約1-2歲時,之後就保持穩定[15] [16] [17] [18] 性早熟症 (precocious puberty)有關。而當青春期來臨時,褪黑素的製造就會減少。在成人時,松果體的石灰化 是典型的。
在動物方面,松果腺在性成熟、冬眠 、新陳代謝以及季節性繁殖上明顯扮演著重要的角色。
松果體的細胞結構 與脊索動物的視網膜細胞似乎有發展的相似性。[7] 鳥類 與爬蟲類 已被發現在其松果體中有黑視素 (melanopsin)這種光傳導 色素 。鳥類的松果體被認為扮演與哺乳動物 的視叉上核 一樣的角色。[19]
針對齧齒動物的研究暗示著松果體可能會影響例如古柯鹼等消遣性毒品[20] 抗憂鬱劑 [21] 神經退化症 (neurodegeneration)[22]
[编辑 ] 形上學與哲學 松果體的分泌活動僅相當地被了解。在歷史上,由於松果體的位置在大腦深處而啟發哲學家們認識到它擁有獨特的重要性。這種聯想使得松果體伴隨著圍繞在其被理解的功能上的神話、迷信與神秘 理論而成了個神秘的腺體。
花了許多時間研究松果體的勒內·笛卡兒 [23] 靈魂 之座」[24] [25]
「我的觀點是這個腺體是靈魂最最重要的座位和我們所有想法形成的地方。我如此認為的理由是我除此之外無法找到大腦的其他部分不是成雙的。既然我們用 一對眼睛來看一件物品、用一雙耳朵來聽一個聲音,而在瞬間從未同時有超過一個想法,這必然是從雙眼或雙耳以及其他地方進來的印象在靈魂細想之前就在身體的 某個部位互相統合的結果。現在整顆頭中除了這個腺體外不可能找到任何這樣的地方了,此外它位於最可能適於這項用途的位置,也就是所有凹面的中央。而且它被 將心靈帶入大腦的頸動脈小支流們所支持及環繞著。」[26]
「松果眼」的概念對於法國作家喬治‧巴塔耶 (Georges Bataille)的生殖哲學來說位居中心地位,此在文學學者丹尼斯‧霍勒(Denis Hollier)其論文《反建築》(Against Architecture)中詳細地被分析。[27] [28] [29]
松果體在加斯東·巴舍拉 的《空間詩學 》(The Poetics of Space)中也被注意到。
海蓮娜‧布拉瓦斯基 (Helena Blavatsky)與愛麗絲‧貝利 (Alice Bailey)等一些最早期的新紀元運動 及神智學 神祕主義者,都寫到了靈魂與和松果體有關的神秘關係。這個限於圈內人的松果體概念直接地在愛麗絲‧貝利 的《白魔法專論 》(A Treatise on White Magic)的一個章節出現。
克卡利‧馬德拉 (Khecarī mudrā)認為松果體對瑜珈姿勢的功能相當關鍵。[30]
在混沌教派 (en:Discordianism )中,松果體是讓一個人與女神厄里斯 交流之處的說法被擁護著。[31]
[编辑 ] 附圖 松果體被歸類在這些圖片中
[编辑 ] 注釋
^ Macchi M, Bruce J. Human pineal physiology and functional significance of melatonin. Front Neuroendocrinol. 2004, 25 (3-4): 177–95. doi :10.1016/j.yfrne.2004.08.001 . PMID 15589268 . ^ Arendt J, Skene DJ. Melatonin as a chronobiotic. Sleep Med Rev. 2005, 9 (1): 25–39. doi :10.1016/j.smrv.2004.05.002 . PMID 15649736 . "Exogenous melatonin has acute sleepiness-inducing and temperature-lowering effects during 'biological daytime', and when suitably timed (it is most effective around dusk and dawn) it will shift the phase of the human circadian clock (sleep, endogenous melatonin, core body temperature, cortisol) to earlier (advance phase shift) or later (delay phase shift) times.". ^ Bocchi G, Valdre G. Physical, chemical, and mineralogical characterization of carbonate-hydroxyapatite concretions of the human pineal gland. J Inorg Biochem. 1993, 49 (3): 209–20. doi :10.1016/0162-0134(93)80006-U . PMID 8381851 . ^ Baconnier S, Lang S, Polomska M, Hilczer B, Berkovic G, Meshulam G. Calcite microcrystals in the pineal gland of the human brain: first physical and chemical studies. Bioelectromagnetics. 2002, 23 (7): 488–95. doi :10.1002/bem.10053 . PMID 12224052 . ^ IngentaConnect High Accumulation of Calcium and Phosphorus in the Pineal Bodies . Ingentaconnect.com. 2006-06-16 [2009-07-06] .^ 6.0 6.1 Luke, Jennifer. Fluoride Deposition in the Aged Human Pineal Gland . Caries Res, 2991 (35): 125–28 [2009-05-20] . ^ 7.0 7.1 Klein D. The 2004 Aschoff/Pittendrigh lecture: Theory of the origin of the pineal gland--a tale of conflict and resolution. J Biol Rhythms. 2004, 19 (4): 264–79. doi :10.1177/0748730404267340 . PMID 15245646 . ^ Science 1967;155(759):220–3. PMID 6015532 ^ 9.0 9.1 Schwab, I. R.; O』Connor, G. R.. The lonely eye (Full text). British Journal of Ophthalmology. March 2005, 89 (3): 256 [2009-02-14] . doi :10.1136/bjo.2004.059105 . ^ Pritchard, Thomas C.; Alloway, Kevin Douglas. Medical Neuroscience (Google books preview). Hayes Barton Press. 1999: 76–77 [2009-02-08] . ISBN 1889325295 . ^ Kurochkin, Evgeny N.; Gareth J. Dyke, Sergei V. Saveliev, Evgeny M. Pervushov, Evgeny V. Popov. A fossil brain from the Cretaceous of European Russia and avian sensory evolution (Full text). Biology Letters. The Royal Society. June 2007, 3 (3): 309–313 [2009-02-14] . doi :10.1098/rsbl.2006.0617 . ^ Lerner AB, Case JD, Takahashi Y. Isolation of melatonin and 5-methoxyindole-3-acetic acid from bovine pineal glands. J Biol Chem. 1960, 235 : 1992–7. PMID 14415935 . ^ Coates, Paul M.. Encyclopedia of Dietary Supplements , Marc R. Blackman, Gordon M. Cragg, Mark Levine, Joel Moss, Jeffrey D. White. CRC Press. 2005: 457 [2009-03-31] . ISBN 0824755049 . ^ Axelrod J. The pineal gland. Endeavour. 1970, 29 (108): 144–8. PMID 4195878 . ^ ^ ^ ^ ^ Natesan A, Geetha L, Zatz M. Rhythm and soul in the avian pineal. Cell Tissue Res. 2002, 309 (1): 35–45. doi :10.1007/s00441-002-0571-6 . PMID 12111535 . ^ Uz T, Akhisaroglu M, Ahmed R, Manev H. The pineal gland is critical for circadian Period1 expression in the striatum and for circadian cocaine sensitization in mice. Neuropsychopharmacology. 2003, 28 (12): 2117–23. doi :10.1038/sj.npp.1300254 . PMID 12865893 . ^ Uz T, Dimitrijevic N, Akhisaroglu M, Imbesi M, Kurtuncu M, Manev H. The pineal gland and anxiogenic-like action of fluoxetine in mice. Neuroreport. 2004, 15 (4): 691–4. doi :10.1097/00001756-200403220-00023 . PMID 15094477 . ^ Manev H, Uz T, Kharlamov A, Joo J. Increased brain damage after stroke or excitotoxic seizures in melatonin-deficient rats. FASEB J. 1996, 10 (13): 1546–51. PMID 8940301 . ^ Descartes and the Pineal Gland (Stanford Encyclopedia of Philosophy )^ Treatise of Man . New York: Prometheus Books; 2003. ISBN 1-59102-090-5 ^ ISBN 978-0-19-514581-6 ^ Descartes and the Pineal Gland (Stanford Encyclopedia of Philosophy )^ Against Architecture: The Writings of Georges Bataille , trans. Betsy Wing, MIT, 1989.^ ^ Visions of Excess: Selected Writings, 1927-1939 (Theory and History of Literature, Vol 14) , trans. Allan Stoekl et al., Manchester University Press, 1985^ Kechari mudra definition by Babylon's free dictionary . Dictionary.babylon.com [2009-07-06] .^ [1] Principia Discordia
[编辑 ] 外部連結
5个分类 :
邊緣系統
维基百科,自由的百科全书
邊緣系統 乃一術語指包含海馬體 及杏仁體 在內,支援多種功能例如情緒 、行為 及長期記憶的大腦 結構。這種被描述為邊緣系統的腦部結構與嗅覺 結構相近[1]
[编辑 ] 構造 邊緣系統包括無數在大腦皮質 及皮質下區域(sub-cortex)的結構。儘管從其引入時其明確角色(exact role)及定義已被多次修訂[2]
此外,以下結構偶爾亦會視為邊緣系統的一部分:
[编辑 ] 相關條目 [编辑 ] 參考 ^ [1] ^ ^ 3.0 3.1 3.2 3.3 3.4 3.5 3.6 3.7 Normandy ^ 4.0 4.1 4.2 4.3 4.4 stanford.edu ^ 5.0 5.1 5.2 5.3 5.4 5.5 5.6 Biology.about.com
2个分类 :
時間生物學
维基百科,自由的百科全书
時間生物學 (英語:Chronobiology ;字首來自希臘語Chronos ,指時間 )又譯生物鐘學 ,廣為人知的生理時鐘 。是一門科學,它的任務是研究生物體內與時間有關的周期性現象,或曰這些現象的時間機制。生物節律是憑經驗總結得出的,但有其生理學和分子生物學基礎。生物鐘學與所謂的生理節律 無關。
[编辑 ] 概述 時間生物學的研究目的,是生物體內生理 和行為 的時間機制。在這種機制中,生物體內部的時鐘系統所產生的節律是主要的。
三大中心問題:
生物節律有哪些類型?它們是怎樣影響生物的生理過程的? 節律是內在的嗎?如果是,哪裡是產生節律的發生器,哪裡是起搏點 ,它們怎麼運作? 哪些是外源性 的,周期性的因素,即是所謂的時間伺服器,它們又是怎樣作用於生物時鐘的? 生物時間機制對所有的生物都很重要,而且在目前所有被研究的生物里科學家都找到了其時間節律現象。生物體內有很多過程雖然彼此相關,但在時間上都是 有所區別的。還有一些過程不但受到內在因素制約,還會受到外界因素影響。時間上的區別之一就是各種行為各具其規律性——在一個大範圍內觀察這種規律性,就 可以稱之為生物節律。周期的長度由毫秒到年不等。細胞分裂,呼吸,心跳和行為只是其中的一些例子。
生物鐘學對人的意義在近年來越來越重要,因為我們的生活越來越頻繁地逆這種生物鐘而行。在醫學方面已經確定,服藥時間對藥效影響甚大。在化療 中,若因就節律給病人服用細胞抑制劑的話,調製藥物的濃度就可以比其他給藥時間所採用的濃度降低很多。
[编辑 ] 生物節律的例子 在下面的表格中列舉了一些人體生理功能的每日周期性變化。 體溫在晚睡醒來之前就已經開始升高。就是說人體已經為快要到來的活動做準備。 就是在黃昏或夜行性 的動物,甚至是植物,都存在這種「做準備的」的現象。 植物在日出之前就會激活光合作用相關器官,為光合作用 做準備,以最長時間的利用光能。 很多植物在日間某些時候會展開或合上其花朵。還有一些植物,在一段日子裡花朵相繼開放,只在特定的鐘點合成香料和花蜜。蟲媒如蜜蜂 就在會恰在此時到訪。
鐘點 (小時) 最高點 2:00 惰性 3:00 出生率 4:00 死亡率 6:00 尿液體積 9:00 睾酮生成 11:00 尿液的酸性 12:00 血蛋白 13:00 健康,體溫 14:00 心跳,麻木狀態 16:00 體重 18:00 血壓 19:00 牙疼 22:00 白血球 24:00 外科手術死亡率
[编辑 ] 生物節律的種類 根據周期長度,將生物節律分為四種:
超晝夜的(亞日的)節律(Infradian Rhythmus),該詞源於拉丁語:「infra」為「底下」,「dies」為「日」,亦即周期比一天長的節律。 例如鳥類的遷徙;季節性的(大概 365.25天長)冬眠 ;還有與退漲潮相關的半月周期 ,如在滿月 、新月 出現大潮,而半月時出現小潮(大概 14.25 天),銀漢魚 只在漲潮時在岸上產卵;或者太陰日節律的,以28.5為周期(磯沙蠶屬 )。 近潮汐節律(Circatidal Rhythmus),跟隨12.5小時的潮汐節律。一些海岸線的動物有這種節律,例如水生的蟹類動物漲潮時才會活動,而生長在岸上的蟹則會在退潮時覓食。 次晝夜(超日)的節律(Ultradian Rhythmus)源於拉丁語的「ultra」(超)和「dies」(天、日),其頻率超過日頻率,就是說一天出現兩次以上(嚴格來說是整數次,這是與近潮汐節律的區別)。這些短於24小時的節律的例子有蝙蝠 的捕食周期、成人90分鐘睡眠循環、垂體 的間歇性荷爾蒙 分泌等。 近晝夜節律(Circadiane Rhythmus)來自拉丁語「circa」(大約)和「dies」,為接近24小時長的節律,如人類睡眠 和蘇醒、植物的葉 運動等。 研究得最徹底的是近晝夜節律,當然有歷史的原因——近晝夜節律比周年節律更明顯,但更重要的是近晝夜節律對人類來說更有現實意義。以下講解若無特別說明,都是指近晝夜節律。
[编辑 ] 時間生物學歷史 在18世紀 天文學家 Jean Jacques d』Ortous de Mairanvon就描述了含羞草 的日間葉運動。通過實驗他得知,即使在黑暗中葉子也會呈現這種節律。類似的報導也見於Georg Christoph Lichtenberg,Christoph Wilhelm Hufeland,林奈 和達爾文 。但直到20世紀 人們才開始對該現象作科學研究。在該領域的先驅有:Wilhelm Pfeffer,Erwin Bünning,卡爾·馮·費舍爾 ,Jürgen Aschoff和Colin Pittendrigh。
對生物節律的一個重要的發現是,很多自然節律在持續的同等強度的實驗室條件下也能產生,就是說生物節律並不依賴於外部環境諸如每日光照和溫度的節律變化。內部時鐘的同步 是通過時間變化的媒介完成的,如光和溫度。
[编辑 ] 生物鐘的位置 最近十幾年的生物鐘研究發現,生物體確實存在日常意義的晝夜「時鐘」,並可以告訴生物體的每日鐘點。它們的晝夜周期的誤差常常可以精確到數分鐘。研 究發現具有晝夜生物鐘性質的組織按照調控機能等級可以分為:中央生物鐘(central clocks)和外周生物鐘(peripheral clocks)。從目前所知道的所有生物鐘模型研究得知,晝夜生物鐘是細胞自主的,也就是說,某些細胞就具有生物鐘的特性。雖然晝夜生物鐘受外界環境(比 如光照溫度等條件)的調控,複雜生物的中央晝夜生物鐘往往並不存在於光感受器上。比如,哺乳動物的中央晝夜生物鐘存在於下丘腦的視交叉上核 (Suprachiasmatic nucleus, SCN)。
從20世紀40年代就已經知道,單細胞生物也有自己的生物鐘。所以從中可得知,生物鐘的運行並不一定需要一個網路作為硬體。 藻類如眼蟲屬 或衣滴蟲 有趨光性 晝夜節律。 草履蟲 有晝夜生理過程。 海生的腰鞭毛蟲 , 如多邊膝溝藻 ,也有自己的晝夜節律。它在日出前一個小時就會浮到水面,形成厚厚的一片,進行光合作用。在有利條件下它們會形成紅潮 。在日落之前它們則會重新潛到海中。晚間它們藉助熒光素酶 發出生物光,人們推測這是可以驅趕天敵撓足亞綱 的。 這些節律也可以在實驗室里通過施加持續的影響而發生。
同時原核生物 (細菌 ,和藍藻 )也有晝夜節律。
直至今天在植物中仍沒找到生物鐘的中央控制部分或是起搏點。現在只能推測,光合作用以及與之聯繫的運動時由遍布植物體的多個時鐘共同控制的。
例如光合作用器官的新陳代謝,在實驗中可以觀察到是由於光照對基因表達 產生影響引起的。 每天在葉綠體 的類囊體 膜上的光收集器(Lhc)都會進行光合作用。光會影響細胞核基因的轉錄 和翻譯 。西紅柿到目前為止已發現19個Lhc-基因。
目前在Lhc-基因的運作機制和其啟動子 方面進行著很多的研究。
在動物中起搏點位於中樞神經系統 。
對於昆蟲 如果蠅存在腦部的腹側的側邊小神經元(Small Ventral Lateral Neurons, sLNv)中,這些神經元表達色素擴散因子(Pigment Dispersing Factor, PDF)。不在光葉中。對黑脈金斑蝶 來說,遷徙時較依靠觸鬚上的生物鐘。[1] 軟體動物 在視網膜 的基底部脊椎動物 在視交叉上核 和松果體 (Pineal organ,Epiphyse)中。松果體分泌褪黑激素 (N-乙醯-5-甲氧基色胺)。
魚 ,兩棲類動物 ,爬行類動物 和很多鳥 類動物中松果體是對光敏感的,除此之外它還控制了除褪黑激素晝夜產生節律外的其他節律,如體溫和進食。從中可得知,松果體比視交叉上核 更早掌管著生物節律。
[编辑 ] 哺乳類動物 哺乳類動物 中松果體 和視交叉 上核共同控制了節律 ,但還有很多其他證據表明,還有其他起搏點 的存在 ,如視網膜 。但這些時鐘 是如何運作 的,還是一個未知數 。
[编辑 ] 人類 如上所述,生物鐘學對人類來說越來越重要。
第一,人類的生活模式越來越偏離生物鐘。輪班制越來越多。第二,人類越來越少去曬太陽。特別在冬天,人類在室內過上大部分的時間,光強度鮮有高於500流明 。在戶外即使是陰天最少有8000流明,而太陽光則有100000流明。因此就生物鐘系統來說人類大多生活在黑暗中。人類的晝夜節律其實每天都需要一次新的「校正」,但現在卻遇上了很大的困難。後果可能是失眠 和飲食失調,精力不足直到深度抑鬱症 。在北歐(如挪威 ),在冬天光效率甚至直逼0。在當地,為治療冬天抑鬱症 人們採取了光療法 。第三,人類越來越頻繁的跨時區 遷移(即從東向西,或從西向東),這是對人類晝夜節律一個重大挑戰。
時間利用的習慣分成兩類。一類晚睡晚起,睡眠 時 間長——「貓頭鷹型」,而「雲雀型」則是早睡早起。這個差別是基因因素引起的,所以要更改是困難的。這也意味著,人類大部分是逆節律生活的。青春期年輕人 幾乎全是貓頭鷹型,因此推遲上課時間一個小時,特別是在冬天,無論對授課效果還是健康都是大有好處的。除了這兩種類型外,還有睡眠時間長短之分。這些類型 可以相互組合。還有一種類型的人,他們對睡眠和日光同步束手無策。
生物鐘學與人類的年齡有關。嬰兒時期次晝夜系統(短的活動時間)和長的睡眠交替,直到晝夜系統 發展到能夠掌管生物鐘為止。但隨著年齡的增長它也會漸漸失效。這也是老年人睡眠和活動障礙的原因。
在一項安排人長時間居住在照不到太陽的地方的實驗裡,發現人類的生理時鐘週期是25小時。
目前的研究顯示,人體中存在一種名為褪黑激素 的賀爾蒙物質,由腦中的松果體 製造產生,一般情況下白天在腦中的濃度較低,在晚上的濃度較高,而且已經證實服用退黑激素萃取物可以使人明顯感受到睡意,所以一般相信褪黑激素是掌控人類生理時鐘的關鍵。有些醫師會建議不要只用安眠藥 調整時差,還要搭配褪黑激素。在夜間避免照射到太多人工光源(如日光燈、LCD螢幕),有助於人體生產褪黑激素,讓生理時鐘配合日照。
[编辑 ] 實驗 如上所述,動物和植物的周期性現象很早就為人所知。1759年 就 有人製作了第一張豆類植物葉運動的近晝夜節律圖表。首先植物的葉子會與槓桿的一端相連,槓桿的另一端放置在一個滾輪之上。若葉子下垂,槓桿會在滾輪上留下 一條向上的線,相反當葉子向上提起的時候,就會得到一條向下的曲線。實驗為期數天。前三天每天光照12小時,第四天起停止光照, 若果這種光是葉運動的原因的話,人們應該會得到這樣的結果,就是葉子在沒有光照的後幾天會一直下垂。但事實並非如此。因此光照並不是葉運動的原因。
20世紀 80年代有實驗,去觀察究竟外在因素會不會產生作用。太空實驗室1號 將真菌 脈孢菌
上世紀最重要的研究手段是基因 的突變 篩選。1970年Konopka首次在黑腹果蠅 (Drosophila melanogaster ) 上應用了這一技術。這種果蠅的成蟲破蛹行為有著明顯的近晝夜節律,接近24小時。就是說蠅破蛹的時刻不是隨機的,而是在一天的特定時刻。若一天已經過了這 一時刻,那麼成蟲不會在當天,而是下一天出蛹。這種節律代代相傳。Konopka找到了三種特變品種並不斷培育其後代:第一種Pershort,並不遵循 這種24小時節律,而是19小時,其後代也如是。第二種Perlong,其周期為29小時。第三種Per-,沒有節律。所有這些特變品種在基因的同一區段 上出現了缺陷。90年代末在不同的哺乳類動物里科學家找到了這些「時鐘基因」(BMal, Clock, MPer1, Mper2, Mper3, Cry1, Cry2)。
20世紀90年代開始,生物鐘學開始了跨學科 協作。該領域的研究不單止著眼於某種方法或是某種現象,而是去尋找其內在的聯繫。微生物學 ,生理學 ,生態學 ,心理學 和數學 為時間生物學提供了重要的支持。而時間生物學的研究對象包括植物和動物,還有人。
時間生物學對畜牧業,社會學和醫學有重要的意義,如輪班制,藥理學 ,精神病學 都離不開時間生物學。行為生理學 研究生物鐘的大腦機制,提供了生理學基礎。
[编辑 ] 參考資料 [编辑 ] 相關條目 [编辑 ] 注釋 [编辑 ] 延伸閱讀 Peter Spork: "Das Uhrwerk der Natur. Chronobiologie - Leben mit der Zeit", Reinbek: Rowohlt Taschenbuch Verlag, (rororo: 61665), 2004, ISBN 3-499-61665-3 Arthur T. Winfree: "Biologische Uhren. Zeitstrukturen des Lebendigen" ISBN 3-925-0887-1 G. Hildebrandt, M. Moser und M. Lehofer: "Chronobiologie und Chronomedizin", Hippokrates Verlag 1998, ISBN 3-7773-1302-5 "Vertebrate Circadian Systems", Structure and Physiology, Ed. J. Aschoff, S. Daan, G.A. Groos, Spinger Verlag ISBN 3-540-11664-8 in englischer Sprache Björn Lemmer: "Chronopharmakologie. Tagesrhythmen und Arzneimittelwirkung." Stuttgart 2004. ISBN 3-8047-1304-1 [编辑 ] 外部連結 [编辑 ] 生物鐘學雜誌 [编辑 ] 其他 Template:神經科學
動作電位
维基百科,自由的百科全书
',又稱' 閾電位靜息電位 狀態的細胞膜 受到適當刺激而產生的,短暫而有特殊波形的,跨膜電位 搏動。細胞產生動作電位的能力被稱為興奮性 ,有這種能力的細胞如神經細胞 和肌細胞 。動作電位是實現神經傳導 和肌肉 收縮的生理基礎。
動作電位的形成與細胞膜上的離子通道 開關相聯繫。一個初始刺激,只要達到了閾電位(而不論超過了多少,這就是全或無定理 ),就能引起一系列離子通道的開放和關閉,而形成離子的流動,改變跨膜電位。而這個跨膜電位的改變尤能引起臨近位置上細胞膜電位的改變,這就使得興奮能沿著一定的路徑傳導下去。
[编辑 ] 動作電位過程的描述 首先細胞膜處於靜息電位 狀態,大概在-90到-70 mV水平。動作電位可分為四個相位:
在初始相 膜電位去極至閾電位 ,這是動作電位產生的前提。刺激可以是旁邊已興奮的細胞膜電位改變,或者是化學突觸 上離子通道中的離子內流。 去極化 過程中,膜電位陡峭上升至正值水平。這個鋒電位 中去極化部分被稱為升支 ,而正的電位值則被稱為超射 。(注意:去極化指的是整個上升過程。)接下來朝靜息電位方向的下降過程被稱為復極化 。 在下降過程中,電位會短時間下降到低於靜息電位水平,然後再上升達到靜息電位,這種靜息電位的增大(絕對值)被稱為超極化 (而下降部分被稱為負後電位,上升部分則是正後電位)。 動作電位持續約1-2 ms(神經元 ),但也可達幾百毫秒 (心臟 )。
動作電位後是不應期 ,這又分為0.5 ms的絕對不應期 和3.5 ms的相對不應期 。前者無論刺激多頻繁多強都不能引起動作電位,而後者則要更強的刺激(閾電位提高了)才能引起動作電位。
[编辑 ] 動作電位的成因 關於靜息電位的形成,請看靜息電位 。
[编辑 ] 離子通道的特徵 細胞膜上有多種離子通道 。而動作電位的產生,則與鈉 和鉀 離子通道有關。這些離子通道的開關狀態與膜電位有關,即是所謂的電壓門控通道 。
例如鈉離子通道,在靜息時它是關閉並且是可激活的 。當去極化到一特定值時就會引起其構象的改變,成為打開 狀態。但是離子通道卻不會持續停留在開放狀態,它會在幾毫秒內關閉。這是通過膜上一蛋白質 的失活域的活動實現的,這個失活域會像塞子一樣堵住離子通道。離子通道這種狀態被稱為關閉並失活的 。過渡狀態關閉但可激活的 只有在完全復極化後才可能出現,而開放可激活 的狀態是在簡單模型中不可能實現的。(文獻中也寫道,一個關閉並失活的 通道在復極過程中首先短時間內還是開放 狀態,然後才改變構象直接成為關閉但可激活的 。再次激活只能發生在完全復極之後,在去極化的細胞膜中不可能存在著過渡狀態開放並失活的 )。
當然,並不是所有的通道在電位到達一定值之時全部打開。更可能的是,通道的處於某種狀態的機率
而狀態之間過渡所需的時間也是因通道而異的。鈉通道從關到開發生在1毫秒之內,而鉀通道則要10毫秒。
除了電壓外,還有其他開關通道的機制。對動作電位來說,有兩種值得一提。一種是與內向整流性鉀通道 Kir 有關,這種通道是不可調控的。但卻有一些帶正電的小分子如精素 ,能夠在去極化到一定程度時堵塞通道孔。另一種機制與鉀通道有關,當細胞間的鈣離子與它結合後會開放。
[编辑 ] 動作電位的過程 [编辑 ] 初始狀態 初始時,細胞膜電位為靜息電位 。鈉通道關閉,只有部分鉀通道(主要是Kir ) 開放,鉀離子決定了靜息電位的大小。而各種離子運動的方向和強度為電化學梯度 所決定。例如鈉離子就有向細胞內擴散的趨向,因為其膜外濃度遠較膜內高。
[编辑 ] 起始相 起始相中,要有一個刺激使得膜電位去極化到達閾值。刺激可由於突觸後離子通道的開放(Na+ , Ca2+ )或是旁邊區域產生的動作電位而形成。
當電位到達- 60 mV時,Kir -通道會被精素阻斷,繼而導致快速的去極化並達到鈉通道開放的閾值。但是外流的鉀離子卻會稍微阻擋這一進程(例如鈣離子依賴的鉀離子通道,由於在去極化過程中鈣離子內流而被激活,因而會稍微阻擋這一進程)。
[编辑 ] 升支和超射 但鈉通道起始要到-60 mV以上電位時才會大量開放。鈉離子懸殊的濃度差,使得鈉離子很快進入細胞內,造成細胞膜的去極化,更多的鈉通道被激活,就有更多鈉離子進入,形成了升支和正位的超射。
[编辑 ] 復極開始 但在電位到達最大正值之前,NaV -通道已經開始失活。然後電壓門控鉀通道開始被激活KV 。兩者雖然閾值相近,但後者開放所需的時間更長,直到去極化達到一定程度才開始顯示其作用。當鈉內流達到最大值時,鉀通道才有一半開放,而當所有鈉通道失活時,鉀通道的開放才到了最大值。所以鈉通道開放最多的時候,是電壓最大值之時,而鉀通道的則位於復極開始後。
[编辑 ] 復極 當復極快至靜息電位時,KV 關閉,Kir (對靜息電位很重要)的堵塞會被去除,NaV 會慢慢激活。
[编辑 ] 後超極化 在很多細胞中(主要是神經元)還可以看到後超極化 。這是因為鉀離子的電導性仍然高於靜息值,鉀離子持續流出,而鈉離子已停止流入,這時電位更接近鉀的平衡電位(見能斯特方程 )。在動作電位時,流入的鈣離子會打開鉀通道,這是導致鉀電導性的增高的原因。當鈣水平下降時,電位才會慢慢恢復正常。工作效率增高的鈉鉀泵對後超極化也有影響。
[编辑 ] 不應期 動作電位後,軸突 在一段時間內不能再響應外界刺激。這個時間段,不應期 的長短,是由NaV 活性恢復所需的時間所決定的。不應期分為「絕對不應期」和「相對不應期」 在動作電位後緊接的絕對不應期 ,當時復極化仍在繼續,通道不能再打開。可以說,此時閾值無限大。
在相對不應期 ,提高刺激強度可以獲得一個弱的動作電位。閾值由無限大向正常值靠近。
[编辑 ] 閾值 動作電位需要電位到達閾值才能發起,此時鈉通道開放,鈉離子內流。科學家嘗試找出這一個閾值,但都以失敗告終。神經元可以在一個範圍的刺激內被激活。因此神經科學家放棄了尋找一特定閾值的努力。從系統理論來說,動作電位的產生是通過被動電位動力學和動作電位動力學的分叉 實現的。但在很多教科書中,當講解到閾值時,只會將它描述為靜息電位和動作電位間的灰色地帶 。
[编辑 ] 特殊的動作電位 除了鈉電壓門控通道外,電壓門控鈣通道也可以引起動作電位(如心肌 和浦肯野細胞 ). 植物細胞也能產生動作電位,例如在植物運動 之時。 [编辑 ] 參見 靜息電位 ,興奮 ,刺激 ,運動終板 ,突觸 ,神經系統 ,神經元 ,心臟 ,不應期 。
[编辑 ] 連接
生理學
神經科學
靜息電位
维基百科,自由的百科全书
靜息電位 (又稱靜態電位 、靜止電位 、靜止膜電位 、休息電位 、休息膜電位 ),是神經細胞處於相對安靜狀態時,細胞膜內外存在的恆定電位差。其主要來源於鈉鉀泵 的活動。靜止膜電位的存在對於神經傳導 而言,是非常重要的。
在神經細胞 未受刺激的狀態,想像一個不會影響細胞的電壓器,將一端電極置於神經細胞膜內,一端置於神經細胞膜外,將可發現細胞膜內外存在一電位差 ,此電位差為70mV(0.07V),這是細胞內相對於細胞外來說,電勢 相差了70mV。
這是由於細胞膜的內外離子 濃度分佈不均所導致的。眾離子中最主要影響的是鉀離子和鈉離子,細胞膜上有多個鈉鉀幫浦 ,它們會進行主動運輸 ,每次把三個鈉離子送到細胞外,把兩個鉀離子送入細胞內,過程中耗用了一個腺苷三磷酸 。 細胞膜上還有鈉離子通道和鉀離子通道,在細胞休息的狀態下,鈉離子通道是完全關閉的,使鈉離子不能進出,而一些鉀離子通道卻會打開,因此若干鉀離子會擴散 出細胞外。(此處是指主動離子通道,事實上膜上在一些被動離子通道,但影響不大。)總體而言,神經細胞內有很多的鉀離子,而細胞外有非常多的鈉離子加上一 些鉀離子,造成外面的正離子比內部的正離子還要多,此即為產生靜止膜電位的主要原因。
[编辑 ] 外部連結
透視 (超感官知覺)
维基百科,自由的百科全书
透視 是一種超感官知覺 。此能力能透過普通感官之外的管道,看到遙遠的人、物體甚至事件,或是能「看透」不透明物,或是能感知到人類正常情況下收不到的能量(如無線電波)。這種訊息在報導中往往描述為和普通眼睛接收的訊息相同(所以叫透視),但也有報導描述為聽覺或肌肉動覺。
透視的英文Clairvoyance 一詞也泛指所有的超感官知覺,不過也許較常指不是得自另一個人的訊息。總之超感官知覺除上述的透視外還有預知 心靈感應
就像所有的超能力 現象一樣,科學界內對此現象有很多爭議。甚至超心理學 界內也有。對於此現象是否存在,以及驗證類似現象的實驗是否有效等等都有爭議。
[编辑 ] 歷史中的透視
介觀物理學
维基百科,自由的百科全书
介觀物理學 是物理學 中一個新的分支學科。「介觀(mesoscopic)」這個詞彙,由Van Kampen於1981年 所創,指得是介乎於微觀和巨觀之間的尺度。介觀物理學所研究的物質 尺度和奈米科技 的研究尺度有很大重合,所以這一領域的研究常被稱為「介觀物理和奈米科技 」。介觀的特徵尺度為:10^-7~-10^-9m。
[编辑 ] 相干尺度和介觀漲落 對於巨觀物質的研究,一般應用統計力學 的方法,考慮大量粒子的平均性質。巨觀系統的尺度遠大於(微觀粒子能夠保持其相干性 的)相干尺度 。在這種情況下,每個系統樣本中各個粒子的運動缺乏關聯,呈現統計 上的無規性 ,系統的整體性質很好的被大量粒子的平均運動所描述:即同一系統的不同樣本性質間的差異很小,所有樣本的性質都由系統的平均值 刻畫,統計漲落 很小。處於介觀尺度的材料,儘管也含有大量粒子,但其系統尺度小於相干尺度,同一樣本中的粒子保持相干運動,各個樣本性質差異極大,系統的平均值不再有效的刻畫系統中所有樣本的性質,或者說存在很大的統計漲落。這種漲落稱之為介觀漲落 ,是介觀材料的一個重要特徵。
[编辑 ] 介觀物理和量子混沌 除了試驗 和技術 上的重要應用外,介觀尺度在理論 上是探索量子混沌 現象的重要場所。混沌 現象是巨觀古典力學 中的普遍現象,但在量子 世界中,目前還不能觀測到低激發態 量子系統的混沌現象。介觀物理研究的物質處於量子體系的高激發態,其微觀性質和對應的巨觀力學性質有很大關聯。對應的巨觀力學系統行為不同的話(可積系統或是混沌系統),材料的微觀性質也會不同。這使得介觀物理成為研究量子混沌以及量子力學 和古典力學過渡關係的重要領域。
[编辑 ] 介觀物理的重要領域 整數量子霍爾效應 、弱定域 、AB環 、分數量子霍爾效應 、量子點 、約瑟夫森結 、NEMS 等。前三者正逐步淡出物理學家的視線。
[编辑 ] 相關條目
時間箭頭
维基百科,自由的百科全书
物理學 在微觀 的層次幾乎完全是時間對稱 的,這意味著物理學定律在時間流逝的方向倒轉之後仍然保持為真。但是在巨觀 層次卻顯得並不是那麼回事:時間存在著明顯的方向。時間箭頭 (又稱時間之箭 )就是用於描述這種不對稱的現象。
所謂「在微觀 的層次幾乎完全是時間對稱 的」,通俗地說意指:隨著尺度的減小,事件逆向發生的機率逐漸趨近於正向發生的機率。當尺度非常小時,我們認為兩者是近似相等的。
[编辑 ] 預備知識 時間對稱性可以通過一個簡單的模擬實現來理解。如果時間是對稱的,你可以將影片的一段鏡頭倒過來放,也能理解到發生的事情。例如引力 是對稱的話你可以將一個行星圍繞太陽運轉的軌道倒過來放,這個路徑仍然符合萬有引力定律 。事實上,因為引力是對稱的,因此太陽系就正是這個樣子。
我們現在想像在月亮 上 面往上扔一個球體的錄像回放。正著放的時候,我們看到球向上移動的過程中逐漸慢下來。如果反過來的話,則球往下掉的速度越來越快。這裡就有點不對稱了,一 個是上升越來越慢,一個是下降越來越快。然而,兩種情況下球都是向著月亮加速(也就是背著月亮的方向減速),因此引力始終還是對稱的。
大多數物理定律都是類似上述這個例子,但是有時間箭頭的情況就不同了。
[编辑 ] 時間箭頭 [编辑 ] 什麼是時間箭頭 廣義地說,時間箭頭是為我們指明之間方向的一些規律。依據這些規律,我們可以明確指出事件發生的先後順序。
例如著名的熵增定理(熱力學第二定律的另一種表述形式)指出,一個封閉系統的熵不會減小。因而我們可以通過測量一個封閉系統的熵來確定時間的方向(包含較大熵的系統有大機率處在時間軸上較"靠後"的位置)。
[编辑 ] 熱力學時間箭頭 熱力學時間箭頭來自熱力學第二定律 ,這一定律認為一個孤立系統 的熵 只能隨著時間流逝不斷增加,而不會減少。熵被認為是無序 的量度,因此第二定律隱含著一種由孤立系統的有序度變化所指定的時間方向(也就是說,隨著時間流逝,系統總是越來越無序)。這種不對稱性可用於區分過去 和未來 。換句話說,孤立系統在未來將越來越無序。
儘管任何孤立系統隨時間流逝越來越無序,系統的各部分卻存在著關聯 。一個簡單的例子是玻璃杯被打碎的過程:最終狀態(碎了的杯子)比初始狀態(完整的杯子)更無序,但在杯子的各部分之間存在關聯——兩塊相鄰碎片的邊緣可以準確吻合。於是,一個孤立系統在過去是有序的且其各部分是無關的,而將來則是無序的但各部分是相關的。
第二定律並不是精確的,任何系統都可能通過漲落到達更低熵的狀態(參見龐加萊各態遍歷定理 )。然而,第二定律描述的是系統向高熵狀態轉化的整體趨勢(統計意義上)。
[编辑 ] 宇宙學時間箭頭 宇宙學時間箭頭 指向宇宙膨脹的方向。它可能跟熱力學箭頭有關,這個箭頭指向一個自由能耗盡後宇宙熱寂 或者高寒的結局。一種另類的觀點是,也許這個箭頭會在引力 的作用下逆轉成大擠壓 ,並在反覆的膨脹-擠壓中進化到適合人類的出現(參見人擇原理 )。
如果這個箭頭與其他時間箭頭有關,那麼未來的方向取決於 整個宇宙是否越來越大。也就是說,宇宙正在膨脹而不是收縮,這只是一個定義的問題。
[编辑 ] 輻射時間箭頭 波,包括無線電波 以及聲波 或甚至扔到水中的石頭激發的水波,都從它們的源頭處向四周擴散,儘管波動方程式 除 了這種形式的擴散波之外,也允許從四周聚攏到中間的收斂波。在很仔細地調整實驗條件後,可以扭轉這個時間箭頭產生收斂波,因此這個時間箭頭可能源於熱力學 箭頭,因為要產生收斂波,需要比產生擴散波更有序地排列波源。因此,自發產生一個收斂波的可能性要比產生一個擴散波小得多。實際上,一個擴散波的傳播往往 會增加熵,而收斂波則會減少熵,而後者是違反熱力學第二定律 的。
[编辑 ] 因果時間箭頭 原因一般被認為發生在結果之前。我們可以控制未來,但無法控制過去。有人認為這並沒有給出一個明確的箭頭。如果熱力學箭頭扭轉,那麼我們會覺得地上 的碎瓷片是原因,而跳進我們手掌的盤子是結果了。事實上,根據第二定律,初始狀態(更有序而更少自相關)比最終狀態總是要簡單些,因此把開始的情況看成兩 件事情中的原因部分總是容易些。
心理學時間箭頭也與因果時間箭頭有關,因為我們總是記得過去同時能夠影響將來(而不是相反),因此我們把過去看成是將來的原因。
因果時間箭頭被認為是熱力學時間箭頭的一種表現形式。
[编辑 ] 弱力時間箭頭 已經證實某些由於弱核力 引起的亞原子反應違反了宇稱對稱性,但這種情況很少發生。根據CPT定理 ,這意味著它們同時是時間不可逆的,由此產生一個時間箭頭。這類過程可能是質子 在早期宇宙產生的原因。
沒有任何機制能說明這個箭頭和其他箭頭有關,如果它反向的話,使我們的宇宙有所區別的就只是那個世界充滿了反質子而不是質子。更準確地說,對質子和反質子的定義將剛好相反。
宇稱的破壞是非常輕微的,這個箭頭只是勉勉強強地指向某一方向。這將它與其他很容易觀察到的時間箭頭區分開來。
[编辑 ] 量子時間箭頭 量子演化由薛丁格方程式 和波函數塌縮 描述,前者是時間對稱的,而後者卻否。波函數塌縮的具體機制還不清楚,因此也不知道這個箭頭與其他的有何關係。儘管在微觀層次,塌縮似乎不會增加或者減少熵,有人相信其中有一種與熱力學箭頭有關的巨觀原理在起作用。根據量子去相干 的理論,如果假定波函數塌縮 只是表面現象,量子力學箭頭就是熱力學箭頭的一個自然結果。
[编辑 ] 心理學時間箭頭 這是人類的經驗中最顯著的箭頭:我們覺得自己似乎正從過去走向未來;我們覺察到並記得過去而不是將來(儘管兩者有時候被認為只是錯覺)。可是物理學 的時間箭頭如何產生這種知覺還不清楚,因為意識的運作太過複雜,時至今日我們仍只有淺薄的了解。也許因果箭頭影響了我們對原因的學習探求過程,從而形成這 種知覺。
也有人認為時間箭頭是在人腦的知覺進化 過 程中受到熱力學第二定律影響的結果,因此心理學箭頭源自熱力學。為了記住事情,我們的頭腦會從一個無序狀態轉變到一個更有序的狀態,或者從一個有序狀態變 成另一個同樣有序的狀態。為了確保新狀態的正確,必須消耗能量,因此便增加了宇宙其餘部分的無序程度。這個無序度上升的程度總是比我們頭腦的有序度增加的 程度大,因此我們就從宇宙的無序度增加過程中記住了事情,我們記住的事情也就總是在過去。
讓我們進一步說明心理學箭頭和熱力學箭頭之間的關係,以便理解。前面提到熱力學第二定律指出在系統不同部分之間的關聯會在將來的方向上逐漸增加 (而不是向過去的方向)。既然記憶就是我們的腦細胞(或者計算機中的二進制數位)和外部世界的聯繫 ,記憶為何應該隨著時間流逝(向著將來方向)增多而不是減少,原因就很明顯了。此外,我們的行為可能影響將來而不是過去,是因為影響外部世界意味著在我們自己(我們的身體或者腦袋)和外部世界之間建立關聯。
[编辑 ] 不可逆性示例 試考慮一個巨大的容器充滿了兩種不同的液體,例如一邊是一種染料而另一邊是水。如果沒有東西在中間分隔這兩種液體,其分子的布朗運動 會導致它們隨著時間推移開始混合在一起。然而,當它們完全混合之後,你不能期待染料和水能自動重新分開。
現在我們重複上面的實驗,但這次我們用一個非常小的、只能容納幾個分子(大概10個)的容器。給定一個相對較短的時間,人們可以想像那些分子會有機會重新分離,所有的染料分子在一邊而水分子在另一邊。正式敘述請參見波動定理 。
對於大點的容器,不能自動分離只是因為這太不可能發生了,甚至耗盡整個宇宙的壽命也不夠。這些液體開始於一個高有序度的狀態,其熵 (有時候這個字定義成「無序」)隨時間增加。
如果從某種較早期的混合狀態開始觀察較大的容器,可能發現它只是部分混合。有足夠的理由認為,如果沒有外部的介入,這些液體目前達到這個狀態是因為過去它更為有序,那時候它的分離度更大,並且在將來則更無序,混合度更高。
[编辑 ] 參見 [编辑 ] 延伸閱讀 胡·普賴斯. 時間之矢與阿基米德之點——物理學時間的新方向 (M). 上海: 上海科學技術出版社. {{{date2001}}}. ISBN 7-5323-5737-6 . Halliwell, J.J. et.al.. Physical Origins of Time Asymmetry. Cambridge. 1994. ISBN 0521568374 . Boltzmann, Ludwig . Lectures On Gas Theory. University Of California Press. 1964.Peierls, R. Surprises in Theoretical Physics. Princeton. 1979. Feynman, Richard . The Character of Physical Law. BBC Publications. 1965.Penrose, Roger . The Emperor's New Mind. Oxford University Press. 1989. ISBN 0198519737 .Penrose, Roger . The Road to Reality. Jonathan Cape. 2004. ISBN 0224044478 .Price, Huw. Time's Arrow and Archimedes' Point. 1996. ISBN 0195100956 . Website Zeh, H. D. The Physical Basis of The Direction of Time. 2001. ISBN 3540420819 . 該書的網頁 柯文尼 譯者:江濤. 時間之箭 (M). 湖南: 湖南科學技術出版社. {{{date2007-6-1}}}. ISBN 9787535715807 . [编辑 ] 外部連結
3个分类 :
時間膨脹
维基百科,自由的百科全书
時間膨脹 是一種物理現象:兩個完全相同的時鐘之中,拿著甲鐘的人會發現乙鐘比自己的走得慢。這現象常被說為是對方的鐘「慢了下來」,但這種描述只會在觀測者的參考系 上才是正確的。任何本地的時間(也就是位於同一個座標繫上的觀測者所測量出的時間)都以同一個速度前進。時間膨脹效應適用於任何解釋時間速度變化的過程。
在阿爾伯特·愛因斯坦 的相對論 中,時間膨脹出現於兩種狀況:
狹義相對論中,時間膨脹效應是相互性的:從任一個時鐘觀測,都是對方的時鐘走慢了(當然我們假定兩者相互的運動的等速均勻的,兩者在觀測對方時都沒有加速度)。
相反,引力時間膨脹卻不是相互性的:塔頂的觀測者覺得地面的時鐘走慢了,而地面的觀測者覺得塔頂的時鐘走快了。引力時間膨脹效應對於每個觀測者都是一樣的,膨脹與引力場的強弱與觀察者所處的位置都有關係。
[编辑 ] 概述 狹義相對論中測定時間膨脹的公式為:
當中
兩個本地事件 (就是在同地方發生的兩個事件)之間的時間間隔——這被稱爲原時 光速 ;而勞侖茲因子 。那麼移動中的那個時鐘走得就比較慢。日常生活中,就算是高速的太空飛行,造成的時間膨脹效應也太小,一般很難被探測到,因此可被忽略。只有在物體達到30,000 km /s(光速的1/10)以上時,時間膨脹才顯得十分重要。
因為勞侖茲因子而引起的時間膨脹現象是於1897年 由Joseph Larmor 發現──最起碼有電子在原子核運轉而引起的現象。
[编辑 ] 實驗證明 時間膨脹的試驗已經做過許多次了。自1950年代開始的粒子加速器 (如歐洲核子研究組織 的加速器)的日常工作,就是持續進行的狹義相對論實驗。具體的幾個實驗包括:
[编辑 ] 速度時間膨脹實驗 Rossi and Hall (1941) 比較了位於山頂和位於海平面的由宇宙射線製造出的渺子 數量。儘管渺子從山頂到地面所需的時間已經是幾個半衰期,但是在海面的渺子數量卻只是少了一點。這是由於渺子相對於測試者以高速運動,導致了可觀的時間膨脹效應。經計算,快速移動的渺子的衰變速度比它們相對測試者靜止時的衰變速度要慢10倍。[编辑 ] 引力時間膨脹實驗 Pound, Rebka 在1959年測量出位於較低海拔(所受重力較強)的光波的頻率有很小的引力紅移 。得出的數值和廣義相對論預測的數值有小於10%的偏差。不久後Pound和Snider在1964年得出更準的1%偏差,正好就是引力時間膨脹預測的效應。[编辑 ] 速度和引力時間膨脹結合實驗 Hafele and Keating 在1971年把兩個銫 原子鐘 分別放在兩架分別向東和西飛的商務客機上,並對比放在美國海軍天文臺 的時鐘。飛機上的原子鐘應該衰變得更快,因爲他們位於距離地面較高,因此引力時間膨脹較小。不過,相反地,它們又會走得較慢,因爲他們相對天文臺的時鐘的速度很快。而當中的引力時間膨脹效應較大,因此兩個時鐘的時間相對走快了。實驗結果和預測的結果相符合。在2005年,英國國家物理實驗室 公佈了他們在另一次相似的實驗中所得出的結果。[1] 倫敦 和華盛頓 ),但是實驗之中的原子鐘更爲精確。公佈的結果誤差為4%。全球定位系統 可被視爲一項持續進行的狹義和廣義相對論實驗。軌道上的時鐘根據時間膨脹效應被調教成適當的速度,以對應位於地面的時鐘。另外,有關廣義相對論的一些微調已經編寫進定位衛星,要不然,每12個小時定位結果便會有大約7米的偏差。[來源請求] [编辑 ] 介子衰期 比較不同速度下μ-介子的衰期是可行的。慢速的介子可在實驗室裏製造,而快速的介子則在宇宙射線穿入大氣層時製造出來。實驗室中靜止介子的衰期為2.22 μs,由宇宙射線製造出來的介子的速度為光速的98%,衰期為比靜止時大5倍左右,和理論相符合。[2]
[编辑 ] 太空飛行與時間膨脹 有了時間膨脹效應,人們在以極高速運動的飛船時,儘管外界已經經歷了很長的歲月,自己卻沒什麼老化,因爲極大的速度會使飛船(和裡面的所有物體)的 時間減慢。也就是說,飛船的時鐘走了一圈,地球上的時鐘已經轉了許多圈了。只要速度夠高,這個效應便會明顯地顯示出來。比方説,旅行者似乎只航行了一年, 對地球上的人來説卻有十年之久。實際上,只要以地球引力加速度(9.80665 m/s2 )行進,旅行者在有生之年完成的旅行距離為光從大爆炸 起到現在走過的距離,也就是137億光年 之遠!這個旅行者回到地球後,地球已經經過了數十億年了。這個題材被用於Pierre Boulle 的長篇小説《人猿星球 》中。
人們更加有可能利用這個效應把人類送到距離我們最近的恆星上,而不需耗掉太空員的一生光陰。然而,要實現這種省時的情況,我們則需要研發一些更新、更先進的推進 技術。另一個問題是,在這麼高的速度下,空間裏的粒子會折射,成爲高能量的宇宙射線 。要想飛船不被毀滅,我們必須用到一些不可思議的防輻射措施。其中一種建議的措施是利用強電磁場把前來的物質離子化 ,或把它們反出去。
目前的太空科技有著許多根本性的限制,如要把飛船加速到接近光速需要大量能量,小型碎片等會對飛船造成威脅。不過,在今天的太空任務中,時間膨脹並 不是考慮的因素之一,因爲就算是頂速也達不到有效的速度。另外一個太空飛行會涉及到的時間膨脹效應情況,是接近一個有著極大引力的地方,如黑洞 ,那裏會有強大的引力時間膨脹效應。
[编辑 ] 以固定加速度前進時的時間膨脹 在狹義相對論中,時間膨脹絕大多數時候都出現在相對運動速度不變的情況下。不過,勞侖茲公式允許我們算出兩物之間有固定加速度 時的時間膨脹,就是一物相對另一個沒有加速度的物體以g 速固定地加速。
設t 為一慣性系統的時間。設x 為空間座標,並設x軸平行於飛船固定加速的路徑。假設飛船的位置是t = 0、x = 0,而其速度為v 0 ,以下的公式有[1] :
位置::
速度:
原時:
作爲x 的函數的靜止系統時間:
[编辑 ] 時間膨脹的簡單推論
相對於A或B的觀測者看到更長的路經,時間 >
2L/c ,速度
c 不變
由於光在任何參照系中的速度都相同,於是時間膨脹可以解釋為下:
想像一個由兩面鏡子A和B組成的時鐘,兩面鏡子相距L ,其間有光束來回反射,光束每次接觸到其中一面鏡子時,時鐘便會走動。
參考系靜止時(右上圖),光來回一次的路徑長度為2L ,時鐘走過的時間為2L 除以光速:
參考系以v 速移動時(右下圖),光的路徑傾斜了,並且更長了。狹義相對論的另一前提是,光在任何參考系中的速度都相同,因此移動中觀測者的時鐘的周期便會加長。這代表,相對於時鐘的參考系中,那個時鐘便顯得走得很慢。簡易的勾股定理 就可導出狹義相對論的預測:
光束走過其路徑的總時間為
路徑一半的距離可寫為已知參量的函數
提出:c Δt '
化簡後
表達了移動時鐘的周期比參考系的時鐘要慢這個事實。
[编辑 ] 時間膨脹在兩個慣性觀測者之間是對稱的 常理會認爲,如果飛船裏的時間被拖慢了,則裡面的太空員會看到外面的世界相對地「加速」了。可是,狹義相對論卻算出相反的結果。
日常生活中其實也有這種怪異的情況:如果兩人相距一段距離,則A會看到B「縮小」了,但是B也覺得A「縮小」了。這種透視 現象已經被人們適應、接受了,因爲它存在於平日的生活裏,但是人們對相對論就毫無準備。
我們已經對有關距離的相對論見解習以爲常了:從北京到上海的距離當然等於上海到北京的距離。另一方面,當我們考慮到速度方面,會認爲如果一個物體在運動,運動一定會是相對於某物:星體、地面或另一人。A物相對B物的速度,是相等於B物相對A物的速度,兩者完全相等。
在狹義相對論中,一個移動中的時鐘相對觀測者的時鐘顯得較慢。如果A和B在不同的飛船上,而相對速度為接近光速,則A(使用自己的時鐘)覺得B時間變慢了,B也覺得A的時間慢了。
注意要在參考系統中建立「同步」的概念,「到底一件事是否和另一處的另一件事同時發生」這個問題有著關鍵的重要性。所有計算都最終要涉及到哪些事件 是同時發生的。也要留意,要建立兩個空間中相隔的事件的同步性,這兩個地方一定要有訊息相互傳遞,這也代表了光速是決定同步性的一個重要因素。
大家當然會問到,狹義相對論怎麼能在A相對B有時間膨脹而B相對A也有時間膨脹的情況下不前後矛盾。要消除矛盾,我們必須丟棄人們日常對同步性的直 覺概念。同步性,是位於一個參考系中的一位觀測者和一系列事件之間的關係。如此類推,我們能接受「左」和「右」是參照於觀測者的位置和方向。這是因爲左和 右是一種物體間的關係。同理,柏拉圖 解釋,「上」和「下」是對應於地球的表面的一種關係,因此人們是不會在他們的對蹠地掉下去。
理論的架構裏有一個同時性的相對論 ,它影響著特定事件如何根據有相對運動的觀測者被調准。由於每個觀測者對兩個事件是否同時發生都有不同的見解(見孿生子佯謬 ),因此任一個觀測者都可認爲對方的時鐘減慢了,這並不會導致理論自相矛盾。這矛盾現象有許多更明確的解釋,如下。
[编辑 ] 時態座標系與時鐘同步 相對論使用時鐘同步的步驟來建立時態座標系。現在這常被稱爲愛因斯坦同步步驟 ,因爲曾出現在他於1905年的論文裏。
一位觀測者發送一束光訊息,根據他的時鐘時間為t 1 。在一處遙遠的事件,這束光被反射回來,在t 2 時到達原先的觀測者(根據同一個時鐘測量)。這個情況下,由於光線來回都以同一個速度走著同一條路線,因此光訊息在遙遠處被彈回來的那一刻的時間t E 為t E = (t 1 + t 2 ) / 2。這樣,使用一個觀測者的一個時鐘便可以定義時態座標系,並在宇宙各處都適用。
對稱的時間膨脹效應發於以這種方式設立的時態座標系中。效應中,另一個時鐘被觀測者認爲走慢了。觀測者並不覺得自己身上發生著時間膨脹,但他可以知道相對另一個座標系,他的時間是顯得較慢。
[编辑 ] 速度時間膨脹的時空幾何
橫向運動的時間膨脹
動畫中的綠點和紅點代表飛船。綠色船隊相互並沒有速度,因此每艘飛船上的時鐘所走的速度都相同,而船隊則可以保持飛船之間的同步。紅色船隊相對綠色船隊移動,速度是光速的0.866倍。
藍點代表光束。根據綠色船隊的時間,光束每來回一次所花時間為2秒,單向所需時間為1秒。
從紅色飛船看(根據自己的時間),兩個紅色飛船之間的光束單向所需時間為1秒。而從綠色飛船來看,紅色飛船之間所發出光線的路徑為一個對角的斜線,單向所需時間為2秒。(以綠色的角度看,紅色飛船每2秒(綠色飛船時間)行進距離為1.73(
其中一艘紅色飛船每秒向綠船發射訊號。根據綠色飛船的時間,每隔2秒才接收一次訊號。動畫中沒有提到的是,所有物理效應都被等比例縮小了。紅色飛船發出的訊號頻率(紅色飛船所測量到的)比綠色飛船接收到的訊號頻率(綠色飛船所測量到的)要高,反之亦然。
此動畫分別以紅色或綠色飛船作爲參考物,藉以強調速度時間膨脹的對稱性質。由於相對論中(牛頓力學 中也如此)沒有絕對運動這回事,因此無論是紅色還是綠色的船隊「在其自己的參考系中」都會認爲自己是不動的。
再次重申,大家要明白,這些交互作用和計算反映了相互運動中飛船的真實狀況。這不是單純的測量或通訊方法中出現的古怪情況。
[编辑 ] 參見 [编辑 ] 參考資料 ^ http://www.npl.co.uk/upload/pdf/metromnia_issue18.pdf ^ JV Stewart. Intermediate electromagnetic theory . Singapore: World Scientific. 2001: p. 705. ISBN 9810244703 . Callender, Craig & Edney, Ralph. Introducing Time. Icon. 2001. ISBN 1-84046-592-1 . Einstein, A. (1905) "Zur Elektrodynamik bewegter Körper", Annalen der Physik , 17 , 891. English translation: On the electrodynamics of moving bodies Einstein, A. (1907) "Über eine Möglichkeit einer Prüfung des Relativitätsprinzips", Annalen der Physik. Hasselkamp, D., Mondry, E. and Scharmann, A. (1979) "Direct Observation of the Transversal Doppler-Shift", Z. Physik A 289 , 151–155 Ives, H. E. and Stilwell, G. R. (1938),「An experimental study of the rate of a moving clock」,J. Opt. Soc. Am , 28 , 215–226 Ives, H. E. and Stilwell, G. R. (1941),「An experimental study of the rate of a moving clock. II」,J. Opt. Soc. Am , 31 , 369–374 Joos, G. (1959) Lehrbuch der Theoretischen Physik , 11. Auflage, Leipzig; Zweites Buch, Sechstes Kapitel, § 4: Bewegte Bezugssysteme in der Akustik. Der Doppler-Effekt . Larmor, J. (1897) "On a dynamical theory of the electric and luminiferous medium", Phil. Trans. Roy. Soc. 190 , 205–300 (third and last in a series of papers with the same name). Poincaré, H. (1900) "La theorie de Lorentz et la Principe de Reaction", Archives Neerlandaies , V , 253–78. Rossi, B and Hall, D. B. Phys. Rev. , 59 , 223 (1941). NIST Two way time transfer for satellites Voigt, W. "Ueber das Doppler'sche princip" Nachrichten von der Königlicher Gesellschaft der Wissenschaften zu Göttingen , 2 , 41–51. [编辑 ] 外部連結
3个分类 :
重力時間延遲效應
维基百科,自由的百科全书
引力時間延遲效應 ,或經常稱作夏皮羅時間延遲效應 是在太陽系 中能夠進行的四個古典廣義相對論 的實驗驗證之一(另外三個是引力紅移 、水星近日點的進動 、光線在太陽引力場中的偏折 )。這種時間延遲效應是指當雷達 信號途徑一個大質量天體時,在觀測者看來這個信號發射到指定目標以及返回的時間都要比沒有大質量天體存在時所需的時間略長。與引力紅移的區別在於它是引力場造成的純粹時間延遲效應,並不改變信號的波長 。
[编辑 ] 歷史 引力時間延遲效應最早由美國 哈佛大學 天體物理學 家歐文·夏皮羅 (Irwin I. Shapiro )於1964年 在理論上提出。1960年 前後,廣義相對論的實驗驗證方法似乎已經被研究殆盡,在當時可研究的內容基本只有在數學層面上的理論研究。1962年 ,理察·費曼 曾 因自己參加的一個引力學術會議失望地發出感慨,他說自己從會議中沒有學到任何東西,引力領域的討論如果不是在糾正前人的錯誤就是毫無用處的內容……自己不 會再去參加任何引力的學術會議了。所幸這種情況在1964年得到了改變:夏皮羅從光線在太陽引力場中偏折這一事實中得到啟發,他認為如果廣義相對論正確, 那麼當光途經太陽引力場時其 速度將會被減緩,減緩量和角度偏移量成正比。夏皮羅同時設想了一個用於證實他的預言的觀測實驗:從地面上向金星 和水星 表面發射雷達波並測量其往返時間。夏皮羅通過計算得到當地球、太陽和金星最大限度地在同一條直線上時,由於太陽質量導致的雷達波往返的時間延遲將達到200毫秒左右,這種延遲量在二十世紀 六十年代 的技術範圍內完全可以觀測到。
位於土星的
卡西尼號 向地球發送的信號在太陽的引力場中延遲
第一次實驗觀測是藉助麻省理工學院 的「草堆」雷達天線(Haystack radar antenna )完成的,其結果和理論預測符合得很好,誤差小於5%。其後這種實驗被不斷重複,並且不斷取得更高的精度。1976年 的海盜號 火星 探測器將精度提高到了0.1%;而2003年 的卡西尼號 土星 探測器的實驗則達到了小於0.002%,是迄今為止精度最高的廣義相對論實驗驗證。
[编辑 ] 時間延遲的計算 根據廣義相對論 理論,原時 和引力場 度規 下的坐標時 存在關係
其中g 00
當光信號途經一個單一質量的引力場時,其時間延遲量為
其中向量 ,而史瓦西半徑 表示,時間延遲量可寫成
這裡R s
[编辑 ] 實例 在行星際探測器(例如旅行者1號 、2號 ,先驅者10號 、11號 )的測距中,由於太陽引力場的作用引力時間延遲效應一定要被考慮到測距的數據中去。而在所有重力波的探測中,來到太陽系的重力波 都會受到太陽、大行星甚至小行星 引力場的影響而產生延遲。特別是在對毫秒脈衝星 的計時觀測中,來自毫秒脈衝星的脈衝信號傳播到地球的時間因受到引力時間延遲的影響會造成偏差。由於對毫秒脈衝星的計時是目前探測超低頻重力波的僅有手段,因而引力時間延遲效應對超低頻重力波的探測的影響也需要被考慮。
[编辑 ] 愛因斯坦關於引力場對光的作用的思考 「其次我們的結果表明,根據廣義相對論理論,光速是一個常數這一定律變得沒有任何佐證。它作為狹義相對論 的 兩條基本假設之一,以及作為我們經常引用的定律,由此將不能保證無限度的正確性。一束光的彎曲只可能是因為光在傳播過程中速度隨位置發生改變而產生。或許 現在我們會猜想是不是狹義相對論,以及它推出的所有相關理論即將灰飛煙滅。但事實並非如此。現在我們只能得出結論:狹義相對論並不能保證無限度的正確性, 它的結論只是在我們能夠忽略引力場對現象(例如,光)的影響時才成立。」——阿爾伯特·愛因斯坦 (《廣義相對論:第22章 - 廣義相對論原理的一些推論》)
[编辑 ] 參見 [编辑 ] 參考文獻 Boost for General Relativity. Nature . 12 July 2001.古騰堡計劃 中收錄的《Relativity : the Special and General Theory by Albert Einstein.》免費電子版本 Irwin I. Shapiro. Fourth Test of General Relativity. Physical Review Letters. December 1964, 13 : 789-791. Irwin I. Shapiro, Gordon H. Pettengill, Michael E. Ash, Melvin L. Stone, William B. Smith, Richard P. Ingalls, and Richard A. Brockelman. Fourth Test of General Relativity: Preliminary Results. Physical Review Letters. May 1968, 20 : 1265–1269. d'Inverno, Ray. Introducing Einstein's Relativity. Oxford: Clarendon Press. 1992. ISBN 0-19-859686-3 . Section 15.6 是面向高年級本科生的有關引力時間延遲效應的介紹Will, Clifford M.. The Confrontation between General Relativity and Experiment. Living Rev. Rel.. 2001, 4 : 4-107. gr-qc/0103036 一個研究生水平的關於廣義相對論實驗驗證的綜述John C. Baez, Emory F. Bunn. The Meaning of Einstein's Equation. Amer. Jour. Phys.. 2005, 73 : 644-652. gr-qc/0103044 Gravity Probe B - Testing Einstein 歷史背景,演示夏皮羅時間延遲的動畫
2个分类 :
分類:超常現象
子分類
本分類包含下列10個子分類,共10個子分類。
本分類包含下列34個頁面,共有34個頁面。
C D E G P T 不 人 分 千 孟 巫 心 歐 比 狐 百 神 羅 聖 藍 貓 陰 靈 順 預 鬼 麥 齊
1个分类 :
超能力
维基百科,自由的百科全书
超能力 ,又稱為特異功能 是指心靈感應 、透視 、預知 、念力 、意影、特殊體質(身體著火、發電)等的超自然能力,被歸類於超心理學 (Parapsychology)的範疇內。
超能力的存在與否始終沒有獲得科學上的真正證實;目前,世界上有超過20個研究機構或組織提供總計超過一百萬美元的獎金(參閱 List of prizes for evidence of the paranormal ),懸賞任何能夠在控制條件下證明超能力真正存在的案例,但是那些獎金至今沒有被任何人取走。
由於超能力的展示,常被詐騙 集團所利用做為斂財的工具。例如像是利用針孔相機 原理,偽裝成透視能力的超能力者。且超能力對於科學驗證基礎的「重現性」,則相當薄弱。因此至今,超能力的相關研究常被視為偽科學 。
再者,在歐洲中古世紀曾以宗教為名,所發起的魔女審判,對於異端能力者的追殺。如果超能力是藉由遺傳取得,則大規模的種族滅絕的行為,就有可能是超能力罕見的原因。
至於後天取得超能力的論點,則認為可以藉助藥物或修練,使腦部活性化,藉以開發潛在能力。
[编辑 ] 概要 超能力主要分為 ESP 和 PK 。
ESP(Extra-sensory perception )稱為超感官知覺 ,泛指辨識一般人所無法認識的現象 、或是指干涉該現象的能力。有接觸感應 (psychometry)、透視 (clairvoyance)、心靈感應 (telepathy)、預知 (precognition)等能力。催眠 (hypnosis)目前已列入心理學 範疇,不是 ESP。
PK(Psychokinesis )稱為念力或念動力,指以意志 力、不碰觸物體 而使物體動作,或是移動粒子以產生高熱、低溫、電磁波 等的能力。其中最具代表性的即為念力 (psychokinesis)、瞬間移動 (teleportation)。
肯定世界 中存在「超能力」的人之中對於「誰能夠使用」也仍存在意見紛歧。
例如:
比較項目 論點1 論點2 誰可以用超能力? 只有少部分的人能使用超能力。 每個人都有超能力潛能,但是會使出超能力效果的人只為少數。 超能力的來源? 先天 性超能力後天 性超能力超能力的取得方式? 可經由潛能學習 開發超能力。 因為意外 事故而得到能力。 超能力的強化? 由訓練可以學習到新的超能力或強化自身的超能力 無法強化能力,例如一名可透視三幅牆壁的人無論怎樣學習也不能讓自己看透第四幅牆壁。 使用超能力時的能量 ? 可經由飲食 、充份休息與睡眠 進行補充。 超能力時的能量是無法補充的消耗品,當用完即無法再使用超能力。
相信特異功能者認為,雖然沒有充份、直接的證據 證明其存在,但有一些間接證據,如宗教 上的神蹟 ,可以佐證特異功能的存在,雖然科學 界也不承認。
林書煌 在《特異致動研究 中值得注意的幾個問題 》一文寫道:「生物 體與周圍環境 存在著多種類型的相互作用存在著能量與信息的交流,包括人體 與生物系統 、化學 系統間的相互作用。其中某些作用的方式與機制暫時不明,稱之為特異現象 。」「我們認為重要的是:經過幾年的徘徊、矛盾與思考,我們日益深刻地體會到,在人體科學研究中必須堅持馬克思主義 的認識論 和方法論 ,以『人天觀』、『整體觀』、『系統論』的思想為指導,將現代 科學技術成就與東方 文化 中的精華結合起來,將氣功 、傳統 醫學 與特異功能的研究結合起來,使之進一步納入科學的軌道,走有自己特色的道路。」[1]
錢穆 則以史書 記載如扁鵲 可隔牆見物等諸多異事,以及孩提時的親眼見證,認為特異功能不可妄加否定其存在的可能性。根據他觀察到諸多特異功能的案例,發現超能力者中,以幼童及女性 佔多數,並指出「西方 科學心 物相異,偏重物質 空間 ,此等事誠屬怪異。中國人 向主心物和合成體,則外物移動,未嘗與心無關。亦可謂此等同屬心理 現象,不必偏向物質上探求。但亦非專屬心理學,須心理物理 混為一體求之,庶可得解。觀此,知此等乃人心之本有功能,亦可謂是人心之自然 功能。及其漸長,多在人事上歷練,則此等功能漸失去。但經特殊訓練,年長後,仍保有此功能者,亦可有之。此等事,中國 社會 常見不鮮。」[2]
在虛構作品 中,常以在少青年時期發揮能力的形象與發展心理學 並談。
在超能力、超常現象關連用語中常見的前綴 「psy-」是源自於希臘語 表示心 、魂 意思的「psyche」。
[编辑 ] 超能力的種類 ESP(超感官知覺 ) PK(念動力) 特殊體質 人體發火(en:Pyrokinesis ) 人體生電(少數人可點亮燈泡或日光燈管) 精神治癒(Spirit Healing)、在不藉助藥物或外科手術等醫療行為的前提下,用手或聲音使病患痊癒,泛稱神蹟 或靈療 [编辑 ] 虛構作品中的超能力 多來自漫畫、動畫、小說、電影中,天馬行空的想像下所繪製來的超能力。
例如:ben 10 、正義聯盟 、神奇四俠 、X戰警 、少年泰坦 、鬼太郎 、閃靈二人組 、學園愛麗絲 、黑之契約者 、超能少女組 、魔法禁書目錄 、科學超電磁砲 、ONE PIECE 、火影忍者 、非凡家庭 、植木的法則 、神不在的星期天 ……等 [编辑 ] 參考
^ ^ 錢穆 《現代中國學術論衡》之〈略論中國心理學 二〉-東大出版社、三民出版社、蘭臺出版社
[编辑 ] 參見



 ~~~~~~~~~~~~~~~
~~~~~~~~~~~~~~~







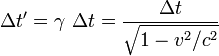
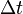 是根據某個觀測者的時鐘,兩個本地事件(就是在同地方發生的兩個事件)之間的時間間隔——這被稱爲
是根據某個觀測者的時鐘,兩個本地事件(就是在同地方發生的兩個事件)之間的時間間隔——這被稱爲 是根據另一個觀測者的時鐘,同兩個事件之間的時間間隔;
是根據另一個觀測者的時鐘,同兩個事件之間的時間間隔;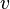 是第二個時鐘相對第一個時鐘移動的速度;
是第二個時鐘相對第一個時鐘移動的速度; 是
是 是
是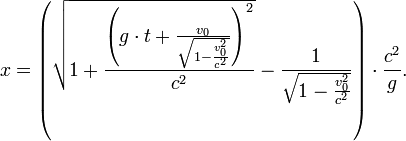
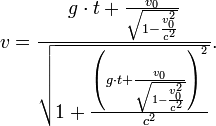
![t^*=\frac{c}{g} \cdot \ln \left[ \left(\sqrt{c^2 + v_0^2} - \frac{v_0}{\sqrt{1-\frac{v_0^2}{c^2}}} \right) \cdot \frac{\sqrt{c^2 + \left(g \cdot t + \frac{v_0}{\sqrt{1-\frac{v_0^2}{c^2}}}\right)^2} + g \cdot t + \frac{v_0}{\sqrt{1-\frac{v_0^2}{c^2}}}}{c^2} \right].](http://upload.wikimedia.org/wikipedia/zh/math/5/0/7/507fe13382e9dfd233e8f0d425469593.png)
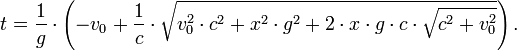




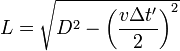

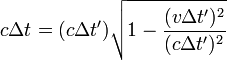


 )光秒。)
)光秒。)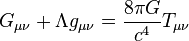


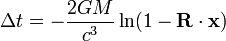
 是觀測者到信號源位置的單位
是觀測者到信號源位置的單位 是觀測者到質量
是觀測者到質量 位置的單位向量。如果用
位置的單位向量。如果用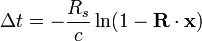
 。
。{kind=link}